Nod-like receptor (NLR) proteins activate pyroptotic cell death and IL-1 driven inflammation by assembling and activating the inflammasome complex. Closely related sensor proteins NLRP1 and CARD8 undergo unique auto-proteolysis-dependent activation and are implicated in auto-inflammatory diseases; however, their mechanisms of activation are not understood.
In this article, the structural basis of how the activating domains (FIINDUPA-CARD) of NLRP1 and CARD8 self-oligomerize to assemble distinct inflammasome complexes is reported. Recombinant FIINDUPA-CARD of NLRP1 forms a two-layered filament, with an inner core of oligomerized CARD surrounded by an outer ring of FIINDUPA.
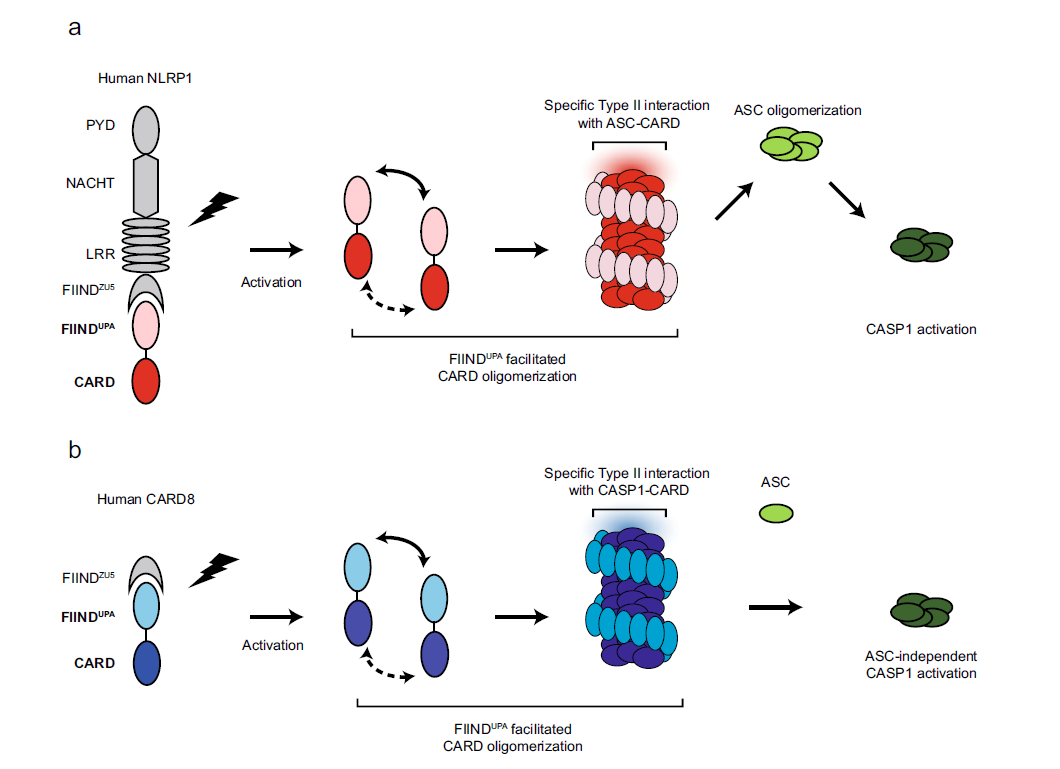
Fig 1: A two-step model for human NLRP1 and CARD8 inflammasome assembly.
From left to right, NLRP1 (a) and CARD8 (b) are “licensed” by auto-proteolysis within the FIIND. Upon activation by PAMP or DAMP triggers, the self-inhibition exerted by the N-terminal proteolytic fragment is relieved. FIINDUPA facilitates the oligomerization of CARD, leading to the formation of a two-layered filament complex. Oligomerized NLRP1-CARD and CARD8-CARD differentially engage ASC or CASP1, leading to the assembly of distinct inflammasome complexes.
Biochemically, self-assembled NLRP1-CARD filaments are sufficient to drive ASC (apoptosis-associated speck-like protein containing a CARD) speck formation in cultured human cells—a process that is greatly enhanced by NLRP1-FIINDUPA which forms oligomers in vitro. The article demonstrates that FIINDUPA and CARD domains play non-redundant roles in NLRP1 activation: NLRP1-CARD is sufficient to form filaments in vitro, trigger ASC speck formation and activate pyroptosis in human cells, albeit with limited efficiency. NLRP1-FIINDUPA does not interact with ASC directly, but dramatically lowers the threshold of NLRP1-CARD oligomerization and filament formation.
Mechanistically, stand-alone FIINDUPA forms ring-like oligomers, whereas recombinant FIINDUPA-CARD forms a two-layered filament, with oligomerized CARD as the inner core around which a FIINDUPA ring spirals. Further analysis reveals that a 9–22-amino-acid linker between FIIND and CARD contributes to the distinct helical symmetry of the NLRP1- and CARD8- FIINDUPA-CARD two-layered filaments. By solving the cryo-EM structures NLRP1- and CARD8-CARD core filaments at 3.7Å, the authors map the precise structural motifs that mediate heterotypic interactions between the CARD domains of NLRP1, CARD8, ASC, and CASP1. This analysis provides a structural explanation as to why activated NLRP1 avidly triggers ASC speck formation, whereas CARD8 directly activates pro-caspase-1. In conjunction with three complementary preprint manuscripts (Articles A-C, See below) published while this manuscript was in revision, the authors’ findings unveil the structural basis for NLRP1 and CARD8 inflammasome assembly and suggest that intrinsic structural properties of the CARD domain can determine the specificity of inflammasome signaling.
Taken together, the article’s cellular, biochemical, and structural findings suggest a model for NLRP1- and CARD8 inflammasome activation (Fig. 1). The article proposes that FIIND auto-cleavage, which has been shown previously to be required for NLRP1 activation, serves as a necessary “licensing” step to liberate the FIINDUPA domain. Upon ligand-induced or 3C protease-mediated activation and relief of auto-inhibition, FIINDUPA self-assembles into ring-like oligomers in a “head-to-tail” fashion (Fig. 2), bringing adjacent CARD monomers into close proximity. This lowers the threshold CARD oligomerization and allows for further “prionoid”-like growth of FIINDUPA-CARD filaments (Fig. 1a). The article’s proposed model of CARD8-FIINDUPA-CARD suggests that this model likely also holds true for the CARD8 inflammasome (Fig. 1b).
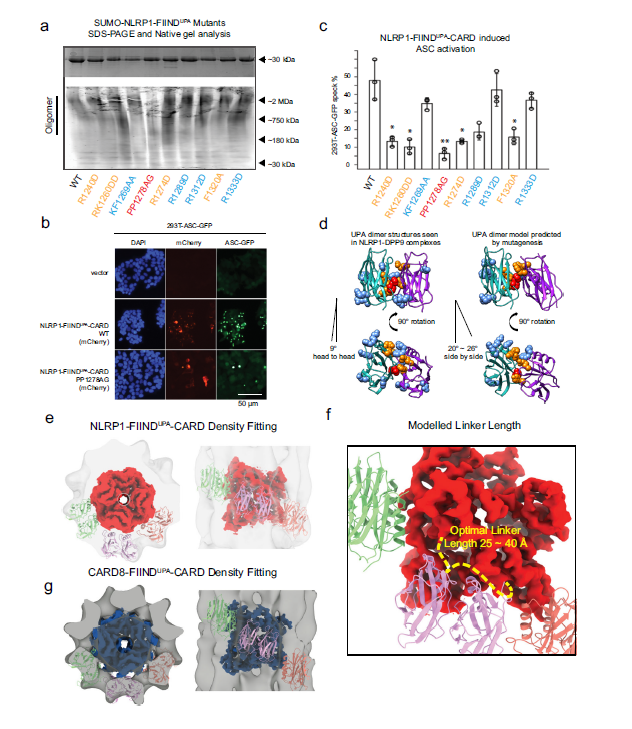
Fig 2. Model of two-layer organization of NLRP1 and CARD8 FIINDUPA-CARD filaments.
(a) Top panel: SDS-PAGE NLRP1-FIINDUPA-CARD mutants. Bottom panel: native gel analysis of SUMO-NLRP1-FIINDUPA constructs, stained by Krypton. (b) ASC-GFP speck formation induced by wild-type NLRP1-FIINDUPA-CARD and PP1278AG in 293T-ASC-GFP cells. (c) Percentage of ASC-GFP specks induced by NLRP1-FIINDUPA-CARD constructs in 293T-ASC-GFP cells. Cells were fixed 24 hours after transfection for fluorescence microscopy. Data are presented as mean values ± SD. P value was calculated with one-way ANOVA, n = 3 biological replicates. ‘*/**/****’ indicates P value < 0.05, 0.01, and 0.0001, correspondingly. (d) Side view and top view of FIINDUPA-dimer structures reported by other groups, and our modeled oligomer interface of NLRP1-FIINDUPA. Potential surface residues that may involve homotypic oligomerization were displayed using spheres, PP1278-1279, were highlighted in red color, R1240, RK1260, R12785, and F1320 are colored in orange. (e) The top and side views of NLRP1-CARD filament and six modeled FIINDUPA domains fitted into the FIINDUPA-CARD filament density. (f) A close-up view of the potential UPA-CARD linker orientation. A linker with 25~40Å is likely to support a regular organization of the two-layer structure. (g) The top and side views of CARD8-CARD filament and si modeled FIINDUPA domains fitted into the CARD8-FIINDUPA-CARD filament density.
The cryo-EM structures of NLRP1-CARD and CARD8-CARD filaments is presented in the article. Despite very similar helical parameters, the subtle differences in rotational angles of individual NLRP1-CARD and CARD8-CARD monomers and their type II junctions give rise to distinct surface charge distributions on the filament scale. Extending this analysis, the authors also found that specific type II interface motifs directly dictate the compatibility of heterotypic pairwise CARD-CARD interactions among NLRP1, CARD8, NLRC4, ASC, and CASP1. In particular, these results explain why NLRP1, but not CARD8, requires ASC for inflammasome activation. Thus, the CARD domains are not mere interchangeable “lego building blocks”. Instead, each possesses intrinsic abilities to discriminate amongst potential downstream binding partners in order to specify the composition of different inflammasome complexes. These distinct inflammasome complexes can likely lead to distinct biological outcomes, in agreement with previous findings demonstrating that different strengths of the pyroptotic signaling are required for cell death and IL-1 cytokine secretion. The prominent role in type II interface-dependent assembly is apparently specific to the inflammasome complexes, as other CARD domain innate immune complexes, such as MAVS (EMD-5925), BCL10 (EMD-7314), RIPK2 (EMD-4399, 6842) rely more heavily on Type I interface junctions, underscoring the importance of studying molecular structures of large complexes at atomic resolutions.
In summary, the findings provide structural insight into the mechanisms of activation for human NLRP1 and CARD8 and reveal how highly specific signaling can be achieved by heterotypic CARD interactions within the inflammasome complexes.
The full article can be accessed here.
A) BioRxiv: Huang, M. et al. Structural and biochemical mechanisms of NLRP1 inhibition by DPP9. 2020.08.13.250241 (2020) https://doi.org/10.1101/2020.08.13.250241.
B) BioRxiv: Hollingsworth, L. R. et al. Novel mode of filament formation in UPA-promoted CARD8 and NLRP1 Inflammasomes. https://doi.org/10.1101/2020.06.27.175497.
C) BioRxiv: Hollingsworth, L. R. et al. DPP9 directly sequesters the NLRP1 Cterminus to repress inflammasome activation. https://doi.org/10.1101/2020.08.14.246132.
1School of Biological Sciences, Nanyang Technological University, Singapore 637551, Singapore. 2NTU Institute of Structural Biology, Nanyang Technological University, Singapore 636921, Singapore. 3Skin Research Institute (SRIS), Agency of Science Technology and Research (A*STAR), 8A Biomedical Grove, #06-06 Immunos, 138648 Singapore, Singapore. 4Lee Kong Chian School of Medicine, Nanyang Technology University, 11 Mandalay Road, 308232 Singapore, Singapore. 5Institute of Molecular and Cell Biology, Agency of Science Technology and Research (A*STAR), 61 Biopolis Dr, 138673Singapore, Singapore. 6Genome Institute of Singapore, Agency of Science Technology and Research (A*STAR), 8A Biomedical Grove, #06-06 Immunos, 138648 Singapore, Singapore. 7Department of Paediatrics, Yong Loo Lin School of Medicine, National University of Singapore, 10 Medical Dr, 117597 Singapore, Singapore. 8The Medical Genetics Department, School of Medicine (KUSoM), Koç University, 34010 Istanbul, Turkey. 9These authors contributed equally: Qin Gong, Kim Robinson.